
Tuesday, September 8, 2009
Monday, August 3, 2009
BER350 Tutorials........
Dear All,
As most of us have classes or practicals on all days of d week........wat i suggest is that we can hav BER350 in the evening at 5pm on ne dy of the week.........do reply which day do you think will be the best for all...........
As most of us have classes or practicals on all days of d week........wat i suggest is that we can hav BER350 in the evening at 5pm on ne dy of the week.........do reply which day do you think will be the best for all...........
Thursday, May 21, 2009
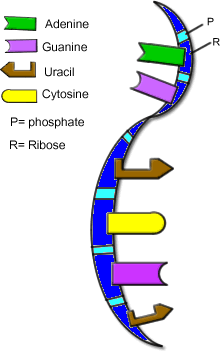
How RNA got started
Scientists may have figured out the chemistry that sparked the beginning of life on Earth.
The new findings map out a series of simple, efficient chemical reactions that could have formed molecules of RNA, a close cousin of DNA, from the basic materials available more than 3.85 billion years ago, researchers report online May 13 in Nature.
Nature.
“This is a very impressive piece of work — a really excellent analysis,” comments chemist James Ferris of the Rensselaer Polytechnic Institute in Troy, N.Y.
The new research lends support to the idea that RNA-based life-forms were the first step toward the evolution of modern life. Called the RNA world hypothesis, the idea was first proposed some 40 years ago. But until now, scientists couldn’t figure out the chemical reactions that created the earliest RNA molecules.
Today, DNA encodes the genetic blueprint for life — excluding some viruses, for those who consider viruses living — and RNA acts as an intermediary in the process, making protein from DNA. But most scientists think it’s unlikely that DNA was the basis of the origin of life, says study coauthor John Sutherland of the University of Manchester in England.
Information-bearing DNA holds the code needed to put proteins together, but at the same time, proteins catalyze the reactions that produce DNA. It’s a chicken-or-egg problem. Scientists don’t think that DNA and proteins could have come about independently — regardless of which came first — and yet still work together in this way.
It’s more plausible that the first life-forms were based on a single molecule that could replicate itself and store genetic information — a molecule such as RNA (SN: 4/7/01, p. 212). RNA world proponents speculate modern DNA and proteins evolved from this RNA-dominated early life, and RNA in cells today is left over from this early time.
While reactions to make RNA from ancient precursors worked on paper, the chemistry didn’t work in the lab. And some scientists thought even RNA molecules were too complex to have spontaneously formed in the primordial soup. Sutherland and his colleagues have shown the reactions are possible.
RNA molecules are formed from three components: a sugar, a base and a phosphate group. In past research, chemists developed each of the components and then tried to put them together to make the complete molecule. “But the components are quite stable, and so they wouldn’t stick together,” Sutherland says. “After 40 years of trying, we decided there had to be a better way of doing this reaction.”
The team took a different approach, starting with a common precursor molecule that had a bit of the sugar and the base. “Basically, we took half a base, added that to half a sugar, added the other piece of base, and so on,” Sutherland says. “The key turned out to be the order that the ingredients are added and the way you put them together — like making a soufflé.”
Another difference is that Sutherland and his team added the phosphate to the mix earlier than in past experiments. Having the phosphate around so early helped the later stages of the reaction happen more quickly and efficiently, the scientists say.
The starting materials and the conditions of the reaction are consistent with models of the geochemistry of an early Earth, the team says.
“But while this is a step forward, it’s not the whole picture,” Ferris points out. “It’s not as simple as putting compounds in a beaker and mixing it up. It’s a series of steps. You still have to stop and purify and then do the next step, and that probably didn’t happen in the ancient world.”
Sutherland and his team can so far make RNA molecules with two different bases, and there are still another two bases to figure out. “It’s related chemistry,” Sutherland says. “That’s how it must have been in the very beginning — a series of fundamental reactions that could make all four types of RNA molecule.”
Once those RNA molecules formed, they would have had to string together to make multiple letters of the code, which could then make proteins. Proteins could then make all the components that make up a cell, and the process would continue from there.
The new findings map out a series of simple, efficient chemical reactions that could have formed molecules of RNA, a close cousin of DNA, from the basic materials available more than 3.85 billion years ago, researchers report online May 13 in
 Nature.
Nature.“This is a very impressive piece of work — a really excellent analysis,” comments chemist James Ferris of the Rensselaer Polytechnic Institute in Troy, N.Y.
The new research lends support to the idea that RNA-based life-forms were the first step toward the evolution of modern life. Called the RNA world hypothesis, the idea was first proposed some 40 years ago. But until now, scientists couldn’t figure out the chemical reactions that created the earliest RNA molecules.
Today, DNA encodes the genetic blueprint for life — excluding some viruses, for those who consider viruses living — and RNA acts as an intermediary in the process, making protein from DNA. But most scientists think it’s unlikely that DNA was the basis of the origin of life, says study coauthor John Sutherland of the University of Manchester in England.
Information-bearing DNA holds the code needed to put proteins together, but at the same time, proteins catalyze the reactions that produce DNA. It’s a chicken-or-egg problem. Scientists don’t think that DNA and proteins could have come about independently — regardless of which came first — and yet still work together in this way.
It’s more plausible that the first life-forms were based on a single molecule that could replicate itself and store genetic information — a molecule such as RNA (SN: 4/7/01, p. 212). RNA world proponents speculate modern DNA and proteins evolved from this RNA-dominated early life, and RNA in cells today is left over from this early time.
While reactions to make RNA from ancient precursors worked on paper, the chemistry didn’t work in the lab. And some scientists thought even RNA molecules were too complex to have spontaneously formed in the primordial soup. Sutherland and his colleagues have shown the reactions are possible.
RNA molecules are formed from three components: a sugar, a base and a phosphate group. In past research, chemists developed each of the components and then tried to put them together to make the complete molecule. “But the components are quite stable, and so they wouldn’t stick together,” Sutherland says. “After 40 years of trying, we decided there had to be a better way of doing this reaction.”
The team took a different approach, starting with a common precursor molecule that had a bit of the sugar and the base. “Basically, we took half a base, added that to half a sugar, added the other piece of base, and so on,” Sutherland says. “The key turned out to be the order that the ingredients are added and the way you put them together — like making a soufflé.”
Another difference is that Sutherland and his team added the phosphate to the mix earlier than in past experiments. Having the phosphate around so early helped the later stages of the reaction happen more quickly and efficiently, the scientists say.
The starting materials and the conditions of the reaction are consistent with models of the geochemistry of an early Earth, the team says.
“But while this is a step forward, it’s not the whole picture,” Ferris points out. “It’s not as simple as putting compounds in a beaker and mixing it up. It’s a series of steps. You still have to stop and purify and then do the next step, and that probably didn’t happen in the ancient world.”
Sutherland and his team can so far make RNA molecules with two different bases, and there are still another two bases to figure out. “It’s related chemistry,” Sutherland says. “That’s how it must have been in the very beginning — a series of fundamental reactions that could make all four types of RNA molecule.”
Once those RNA molecules formed, they would have had to string together to make multiple letters of the code, which could then make proteins. Proteins could then make all the components that make up a cell, and the process would continue from there.
Saturday, April 25, 2009
Researchers Reveal Complete Genetic Sequence of Cow
By Jessica Berman, Washington, 24 April, 2009
An international consortium of researchers has completed mapping the genetic blueprint of the domestic cow, a source of nutrition and livelihood for billions of people around the world.
 Scientists say the landmark accomplishment will lead to better food production and improvements in human medicine.
Scientists say the landmark accomplishment will lead to better food production and improvements in human medicine.The completed bovine genome map is the first full sequencing of a mammalian livestock species. It provides new insight into what makes a cow a cow and identifies important genetic traits that distinguish it from humans and other mammals.
The map, published Friday in the journal Science, is the fruit of a six-year international research effort called the Bovine Genome Sequencing and Analysis Project. The consortium of 300 international scientists mapped the nearly 22,000 genes, or hereditary building blocks, of a Hereford cow - a beef-producing ruminant in wide use around the world.
Scientists compared the cow genome to that of seven other mammals - including the human, dog, rat, mouse, opossum and platypus - and found they share a core set of more than 14,000 genes.
Humans share 80 percent of their genetic sequence with cows, according to the scientists, who say we're more closely related to bovines than to rats and mice.
Researcher Kim Worley at the Baylor College of Medicine in Houston, Texas, which spearheaded the research, says the completed map of the cow genome has implications for laboratory research into new drugs.
"What that means is that when we want to study something that's a human protein we might get better information by studying it in cattle than in mice and rats," Worley said.
Jane Peterson of the Human Genome Project at the US National Institutes of Health, which helped fund the consortium's research, says the cow has been a model for studying human hormone disorders. She says having the entire gene sequence available for study makes the cow a more useful research tool.
"Bovine insulin, for example, was used to find the composition of insulin. And it's also been a model for studying many of the endocrine diseases and the conditions in female cows that are directly related to human[s]," she said.
Cracking the bovine genetic code will also make it a lot simpler and cheaper for cattle breeders to enhance the desirable traits in cattle to produce higher-quality milk and meat, according to geneticist Curt Van Tassell, with the U.S. Department of Agriculture in Beltsville, Maryland.
Van Tassel led a team comparing the bovine sequence in the Hereford cow to six other breeds of cattle, studying important variations, and focusing on the special genes governing cattle reproduction, digestion, lactation and disease resistance.
He says breeders will no longer have to wait years for a bull to produce offspring to learn whether they have desirable genes.
"Specifically what we're able to do is generate a genetic value for a bull the day he's born essentially ... that replaces a process that took six to seven years and $25,000 to $50,000 by collecting performance data on daughters," he said.
A commentary in Science magazine hails the landmark genetics work, predicting that the bovine genetic roadmap may lead to many other important spin-offs.
Wednesday, April 15, 2009
bel302
Prof Narang has asked us to submit the assignment by monday.
The chapter is available at scoops.
Minor 2 copies r checked so contact Prof Gomes. He will show d copies in a day or 2
The chapter is available at scoops.
Minor 2 copies r checked so contact Prof Gomes. He will show d copies in a day or 2
Wednesday, April 8, 2009
Open House
Mail by Dr. D Sundar :
Dear All,
As you may be aware, the IIT-D 'Open House' is scheduled for April 18
(saturday). I would be coordinating the activities for our Department and
would like to have some of you as Activity Heads/Volunteers. It is a day
long affair from 9-5 pm.
Please pass on this message to your other batchmates and seniors/juniors
who may be interested. I look forward to active partcipation from your
side.
Regards,
-Sundar
Dear All,
As you may be aware, the IIT-D 'Open House' is scheduled for April 18
(saturday). I would be coordinating the activities for our Department and
would like to have some of you as Activity Heads/Volunteers. It is a day
long affair from 9-5 pm.
Please pass on this message to your other batchmates and seniors/juniors
who may be interested. I look forward to active partcipation from your
side.
Regards,
-Sundar
Monday, April 6, 2009
Evolution in a bottle

One of the most important experiments in evolution is going on right now in a laboratory in Michigan State University. A dozen flasks full of E. coli are sloshing around on a gently rocking table. The bacteria in those flasks has been evolving since 1988--for over 44,000 generations. And because they've been so carefully observed all that time, they've revealed some important lessons about how evolution works.
The experiment was launched by MSU biologist Richard Lenski. I wrote about Lenski's work last year in the New York Times, and in more detail my new book Microcosm. Lenski started off with a single microbe. It divided a few times into identical clones, from which Lenski started 12 colonies. He kept each of these 12 lines in its own flask. Each day he and his colleagues provided the bacteria with a little glucose, which was gobbled up by the afternoon. The next morning, the scientists took a small sample from each flask and put it in a new one with fresh glucose. And on and on and on, for 20 years and running.
The experiment was launched by MSU biologist Richard Lenski. I wrote about Lenski's work last year in the New York Times, and in more detail my new book Microcosm. Lenski started off with a single microbe. It divided a few times into identical clones, from which Lenski started 12 colonies. He kept each of these 12 lines in its own flask. Each day he and his colleagues provided the bacteria with a little glucose, which was gobbled up by the afternoon. The next morning, the scientists took a small sample from each flask and put it in a new one with fresh glucose. And on and on and on, for 20 years and running.
Based on what scientists already knew about evolution, Lenski expected that the bacteria would experience natural selection in their new environment. In each generation, some of the microbes would mutate. Most of the mutations would be harmful, killing the bacteria or making them grow more slowly. Others would be beneficial allowing them to breed faster in their new environment. They would gradually dominate the population, only to be replaced when a new mutation arose to produce an even fitter sort of microbe.
Lenski used a simple but elegant method to find out if this would happen. He froze some of the original bacteria in each line, and then froze bacteria every 500 generations. Whenever he was so inclined, he could go back into this fossil record and thaw out some bacteria, bringing them back to life. By putting the newest bacteria in his lines in a flask along with their ancestors, for example, he could compare how well the bacteria had adapted to the environment he had created.
Over the generations, in fits and starts, the bacteria did indeed evolve into faster breeders. The bacteria in the flasks today breed 75% faster on average than their original ancestor. Lenski and his colleagues have pinpointed some of the genes that have evolved along the way; in some cases, for example, the same gene has changed in almost every line, but it has mutated in a different spot in each case. Lenski and his colleagues have also shown how natural selection has demanded trade-offs from the bacteria; while they grow faster on a meager diet of glucose, they've gotten worse at feeding on some other kinds of sugars.
Last year Lenski was elected to the National Academy of Sciences. This week he is publishing an inaugural paper in the Proceedings of the National Academy of Sciences with his student Zachary Blount and postdoc Christina Borland. Lenski told me about the discovery behind the paper when I first met him a few years ago. He was clearly excited, but he wasn't ready to go public. There were still a lot of tests to run to understand exactly what had happened to the bacteria.
Now they're sure. Out of the blue, their bacteria had abandoned Lenski's their glucose-only diet and had evolved a new way to eat.
After 33,127 generations Lenski and his students noticed something strange in one of the colonies. The flask started to turn cloudy. This happens sometimes when contaminating bacteria slip into a flask and start feeding on a compound in the broth known as citrate. Citrate is made up of carbon, hydrogen, and oxygen; it's essentially the same as the citric acid that makes lemons tart. Our own cells produce citrate in the long chain of chemical reactions that lets us draw energy from food. Many species of bacteria can eat citrate, but in an oxygen-rich environment like Lenski's lab, E. coli can't. The problem is that the bacteria can't pull the molecule in through their membranes. In fact, their failure has long been one of the defining hallmarks of E. coli as a species.
If citrate-eating bacteria invade the flasks, however, they can feast on the abundant citrate, and their exploding population turns the flask cloudy. This has only happened rarely in Lenski's experiment, and when it does, he and his colleagues throw out the flask and start the line again from its most recently frozen ancestors.
But in one remarkable case, however, they discovered that a flask had turned cloudy without any contamination. It was E. coli chowing down on the citrate. The researchers found that when they put the bacteria in pure citrate, the microbes could thrive on it as their sole source of carbon.
In nature, there have been a few reports of E. coli that can feed on citrate. But these oddballs all acquired a ring of DNA called a plasmid from some other species of bacteria. Lenski selected a strain of E. coli for his experiments that doesn't have any plasmids, there were no other bacteria in the experiment, and the evolved bacteria remain plasmid-free. So the only explanation was that this one line of E. coli had evolved the ability to eat citrate on its own.
Blount took on the job of figuring out what happened. He first tried to figure out when it happened. He went back through the ancestral stocks to see if they included any citrate-eaters. For the first 31,000 generations, he could find none. Then, in generation 31,500, they made up 0.5% of the population. Their population rose to 19% in the next 1000 generations, but then they nearly vanished at generation 33,000. But in the next 120 generations or so, the citrate-eaters went berserk, coming to dominate the population.
This rise and fall and rise suggests that the evolution of citrate-eating was not a one-mutation affair. The first mutation (or mutations) allowed the bacteria to eat citrate, but they were outcompeted by some glucose-eating mutants that still had the upper hand. Only after they mutated further did their citrate-eating become a recipe for success.
The scientists wondered if other lines of E. coli carried some of these invisible populations of weak citrate-eaters. They didn't. This was quite remarkable. As I said earlier, Lenski's research has shown that in many ways, evolution is repeatable. The 12 lines tend to evolve in the same direction. (They even tend to get plump, for reasons yet to be understood.) Often these parallel changes are the result of changes to the same genes. And yet when it comes to citrate-eating, evolution seems to have produced a fluke.
To gauge the flukiness of the citrate-eaters, Blount and Lenski replayed evolution. They grew new populations from 12 time points in the 33,000-generations of pre-citrate-eating bacteria. They let the bacteria evolve for thousands of generations, monitoring them for any signs of citrate-eating. They then transferred the bacteria to Petri dishes with nothing but citrate to eat. All told, they tested 40 trillion cells. Here's a movie of what that looks like...
Out of that staggering hoard of bacteria, only a handful of citrate-eating mutants arose. None of the original ancestors or early predecessors gave rise to citrate-eaters; only later stages in the line could--mostly from 27,000 generations or beyond. Still, even among these later E. coli, the odds of evolving into a citrate-eater was staggeringly low, on the order of one-in-a-trillion.
Now the scientists must determine the precise genetic steps these bacteria took to evolve from glucose-eaters to citrate-eaters. In order to eat a particular molecule, E. coli needs a special channel in its membranes through which to draw it. It's possible, for example, that a channel dedicated to some other molecule mutated into a form that could also take in citrate. Later mutations could have fine-tuned it so that it could suck in citrate quickly.
If E. coli is defined as a species that can't eat citrate, does that mean that Lenski's team has witnessed the origin of a new species? The question is actually murkier than it seems, because the traditional concept of species doesn't fit bacteria very comfortably. (For the details, check out my new article on Scientific American, "What is a Species?") In nature, E. coli swaps lots of genes with other species. In just the past 15 years or so, for example, one disease-causing strain of E. coli acquired hundreds of genes not found in closely related E. coli strains. (See my recent article in Slate.) Another hallmark of E. coli is its ability to break down lactose, the sugar in milk. But several strains have lost the ability to break it down. (In fact, these strains were originally given a different name--Shigella--until scientists realized that they were just weird strains of E. coli.)
Nevertheless, Lenski and his colleagues have witnessed a significant change. And their new paper makes clear that just because the odds of such a significant change are incredibly rare doesn't mean that it can't happen. Natural selection, in fact, ensures that sometimes it does. And, finally, it demonstrates that after twenty years, Lenski's invisible dynasty still has some surprises in store.
Source: Z.D. Blount, C.Z. Borland, and R.E. Lenski, "HI istorical Contigency and the Evolution of a Key Innovation in an Experimental Population of Escherichia coli." PNAS in press (http://www.pnas.org/cgi/doi/10.1073/pnas.0803151105) [UPDATE: PDF AVAILABLE ON LENSKI'S SITE.]
Now the scientists must determine the precise genetic steps these bacteria took to evolve from glucose-eaters to citrate-eaters. In order to eat a particular molecule, E. coli needs a special channel in its membranes through which to draw it. It's possible, for example, that a channel dedicated to some other molecule mutated into a form that could also take in citrate. Later mutations could have fine-tuned it so that it could suck in citrate quickly.
If E. coli is defined as a species that can't eat citrate, does that mean that Lenski's team has witnessed the origin of a new species? The question is actually murkier than it seems, because the traditional concept of species doesn't fit bacteria very comfortably. (For the details, check out my new article on Scientific American, "What is a Species?") In nature, E. coli swaps lots of genes with other species. In just the past 15 years or so, for example, one disease-causing strain of E. coli acquired hundreds of genes not found in closely related E. coli strains. (See my recent article in Slate.) Another hallmark of E. coli is its ability to break down lactose, the sugar in milk. But several strains have lost the ability to break it down. (In fact, these strains were originally given a different name--Shigella--until scientists realized that they were just weird strains of E. coli.)
Nevertheless, Lenski and his colleagues have witnessed a significant change. And their new paper makes clear that just because the odds of such a significant change are incredibly rare doesn't mean that it can't happen. Natural selection, in fact, ensures that sometimes it does. And, finally, it demonstrates that after twenty years, Lenski's invisible dynasty still has some surprises in store.
Source: Z.D. Blount, C.Z. Borland, and R.E. Lenski, "HI istorical Contigency and the Evolution of a Key Innovation in an Experimental Population of Escherichia coli." PNAS in press (http://www.pnas.org/cgi/doi/10.1073/pnas.0803151105) [UPDATE: PDF AVAILABLE ON LENSKI'S SITE.]
Friday, April 3, 2009
change of slots
hey we r doing a course bel301 in c slot
plz check d possibility of replacing to h slot,,
we can get gud oc in c slot
plz check d possibility of replacing to h slot,,
we can get gud oc in c slot
change of slot
hiya people
we have a course chl103 in the b slot
so some of us were thinking that if we could shift it to h slot (the one for genetics this sem) it would be better since we have some good oc's in b slot.
so if you guy's agree we could ask toffee to talk to the authorities.
we have a course chl103 in the b slot
so some of us were thinking that if we could shift it to h slot (the one for genetics this sem) it would be better since we have some good oc's in b slot.
so if you guy's agree we could ask toffee to talk to the authorities.
Wednesday, April 1, 2009
Confidence - It is no more beauty than yourself
How could we become a little more confident especially when we need it, even though , that this can take courage and determination if it is not part of our personality and when this is so, have you noticed we rarely put ourselves into a situation that could make us a bit unsure of ourselves? There are many times, because of uncertainty, we miss out on something that could have been a learning process-perhaps the very thing that would have helped us into having more confidence, self assurance, trust and faith in ourselves.
It may take courage and determination to become confident yet interestingly enough, the people who appear to have this nature are not always confident in every situation; it must be very rare to find anyone who is. Whether or not we are born with confidence, the development of it can have a lot to do with the way one is nurtured from birth-yet this quality can get lost at some stage of growth due to the effects of the distruptive life events being thrown up at us from time to time. Some people gain self confidence with age, whilst others lose it as they get older, yet whatever influence has been the cause of having none or very little confidence doesn’t mean that it is not accessible , because there are a few skills that can help us to get more self assured.
Embrace your qualities
If you have a problem with this , think about some other person who you respect and admire then write down all the qualities you see in them. Now look at the list-you may be surprised to find that all those qualities didn’t come from the people you visualized, those thoughts of qualities you had were created from the feelings deep within you. Every quality you named in thought, whether it was of confidence, self assurance or anything else was brought to the surface from what was already within you; if this weren’t the case how could you recognize them?
How do we see good?
- When we see beauty remember, that it is no more beauty of your self.
- Beauty is reflected in the objects and in the observers who receive beauty from outside. The fact that beauty is seen proves that there is beauty already present in the being of the observer". - Plato
And so whatever we see in other people or whatever we think about anything external comes from within us. The quotation of Plato doesn’t mean that the beauty he speaks of is a physical nature; a person who is not so beautiful on the outside can be very beautiful on the inside. For example , you may have experienced the times you have bought a funny looking-shaped potato, carrot or apple and yet have found that hey tasted really delicious.Who said we have to look perfect ? Who is perfect anyway?
Thoughts and Reactions
If we weigh up the merits of this, then the quality of confidence is already there- may be as said earlier, it just got lost over the years. Being aware of the link between thoughts and reactions may help us not to let it rule any decisions we make which may affect our confidence. Three little suggestions may help to dissolve a reaction before it starts and they are-Listen. Pause. Respond. Remembering to do this help to ward off any unnecessary negative reactions.
We should also be aware of the fact , that for every quality we have, there is an opposite, that is, good or bad, negative and positive, strength and weaknesses. Which of these we come from depends a lot on , which of these we are going to create and nuture , for whatever it is we need to become, or maybe assist us in moving towards bringing up-to-date with the lost or forgotten confidence.
We can not always make ourselves be something that we are not, yet we can pretend. There is an expression "Fake it until you make it "- keep on practicing this and if there are any negative thoughts telling you that you can’t just listen but don’t act on them, tell yourself that you can, (I CAN by way has two meanings i.e. a) an ability and b)an opportunity. Here too is another bit of wisdom that says:-
If you think you are beaten you are .
If you think you dare not , you don’t.
If you’d like to win but think you can’t , it’s almost certain you won’t.
If you think you’ll lose, you’re lost.
The 'IFS' and 'BUTS' keep us from being Confidence
We should also be aware of our thoughts, they are very powerful, they can create either negative or positive issues yet we can change that just by changing those thoughts.take notice of people who believe to be confident, listen to a speaker who appears to be self assured and use them to imitate and practice on. There may be occasions when a lecturer asks a question- you know the answer and would love to say something yet keeping quiet is easier instead of pushing through because that what you wanted to say wasn’t clever or eloquent enough and yet pushing through that barrier in spite of feeling nervous, will get easier to do it again and with gained confidence. It doesn’t matter whether or not you an experienced speaker; what you have to say could be a pearl of wisdom to someone.
However afraid one is to speak-up, in whatever the circumstances are, it is never worse than the few seconds thinking about doing whatever it is that is causing the nervousness or the self-consciousness. If you find that your thoughts are stopping you and are unable to change them then stay with the thought, say to yourself "Yes I am nervous" this is getting you to acknowledge the thought, in other words you have taken possession of the feeling instead of the opposite happening and any nervousness will ease. The more you fight it the worse it gets. Usually it is all the ‘ifs’ and ‘buts’ keep us from being confident.
Breathing controls the Mind
Breathing is really important too , if you breathe from the throat using shallow breathing, this will affect the throat and it begins to dry- this may cause you to panic. The way we breathe also has a bearing on controlling the mind. Remember that the mind effects how we react but the breathing controls the mind. Taking some deep breaths now and again will help you to ease any nervousness. Getting into the practice of exhaling negative and inhaling positive can be a good way to start each day. If , in any situation you are nervous, more than likely your voice will be high pitched and this will convey nervousness or even uncertainty to the listener, lowering your voice indicates confidence. Begin by having trust and belief in what you are saying this will help to make you more confident. Clear communication grabs attention. When speaking to an audience projecting the voice is important, there is nothing more irritating than not being able to hear what the speaker is saying.
Just be Yourself
If there are still some doubts in spite of watching others speak and the little practice of ‘Faking it’ etc. , as well as the breath control programme, start from where you are now, as said earlier change may affected you , i.e. situations etc. even illness puts us in touch with how vulnerable we are. Give yourself permission to be who you are and whatever you are. A lot of the time we don’t do this and then there is the possibility of undermining oneself, this can be an obstruction that we are perfect just the way we are can be instrumental in seeking confidence within ourselves- then will find it was there all the time!
Sunday, March 29, 2009
Attitude is Everything
The process of human change begins within us. We all have tremendous potential. We all desire good results from our efforts. Most of us are willing to work hard and to pay the price that success and happiness demand.
Each of us has the ability to put our unique human potential into action and to acquire a desired result. But the one thing that determines the level of our potential, that produces the intensity of our activity, and that predicts the quality of the result we receive is our attitude.
Attitude determines how much of the future we are allowed to see. It decides the size of our dreams and influences our determination when we are faced with new challenges. No other person on earth has dominion over our attitude. People can affect our attitude by teaching us poor thinking habits or unintentionally misinforming us or providing us with negative sources of influence, but no one can control our attitude unless we voluntarily surrender that control.
No one else "makes us angry." We make ourselves angry when we surrender control of our attitude. What someone else may have done is irrelevant. We choose, not they. They merely put our attitude to a test. If we select a volatile attitude by becoming hostile, angry, jealous or suspicious, then we have failed the test. If we condemn ourselves by believing that we are unworthy, then again, we have failed the test.
If we care at all about ourselves, then we must accept full responsibility for our own feelings. We must learn to guard against those feelings that have the capacity to lead our attitude down the wrong path and to strengthen those feelings that can lead us confidently into a better future.
If we want to receive the rewards the future holds in trust for us, then we must exercise the most important choice given to us as members of the human race by maintaining total dominion over our attitude. Our attitude is an asset, a treasure of great value, which must be protected accordingly. Beware of the vandals and thieves among us who would injure our positive attitude or seek to steal it away.
Having the right attitude is one of the basics that success requires. The combination of a sound personal philosophy and a positive attitude about ourselves and the world around us gives us an inner strength and a firm resolve that influences all the other areas of our existence.
Each of us has the ability to put our unique human potential into action and to acquire a desired result. But the one thing that determines the level of our potential, that produces the intensity of our activity, and that predicts the quality of the result we receive is our attitude.
Attitude determines how much of the future we are allowed to see. It decides the size of our dreams and influences our determination when we are faced with new challenges. No other person on earth has dominion over our attitude. People can affect our attitude by teaching us poor thinking habits or unintentionally misinforming us or providing us with negative sources of influence, but no one can control our attitude unless we voluntarily surrender that control.
No one else "makes us angry." We make ourselves angry when we surrender control of our attitude. What someone else may have done is irrelevant. We choose, not they. They merely put our attitude to a test. If we select a volatile attitude by becoming hostile, angry, jealous or suspicious, then we have failed the test. If we condemn ourselves by believing that we are unworthy, then again, we have failed the test.
If we care at all about ourselves, then we must accept full responsibility for our own feelings. We must learn to guard against those feelings that have the capacity to lead our attitude down the wrong path and to strengthen those feelings that can lead us confidently into a better future.
If we want to receive the rewards the future holds in trust for us, then we must exercise the most important choice given to us as members of the human race by maintaining total dominion over our attitude. Our attitude is an asset, a treasure of great value, which must be protected accordingly. Beware of the vandals and thieves among us who would injure our positive attitude or seek to steal it away.
Having the right attitude is one of the basics that success requires. The combination of a sound personal philosophy and a positive attitude about ourselves and the world around us gives us an inner strength and a firm resolve that influences all the other areas of our existence.
Saturday, March 21, 2009
Solutions to Assignment 3 - BEL204
Clue to the assignment no.3
1. In these studies always look for the supplement that is + for all the mutants. In this case G. So, G is the final metabolite. Then work backward, like supplement that gives ++ and so on.
Sequence is
5 4 2 1 3
E -------- A -------- C--------- B -------- D ---------- G
2. Here, either trpB or trp A is closer to cys
In the first case, recipient is cys+trpA-. So, it may be cys+ trpA- trpB+ or cys+trpB+ trpA- and incoming DNA fragment is cys- trpA+ trpB- or cys- trpB- trpA+. In either case a double crossover event will form prototroph (all +ve).
In the second case, recipient is cys-trpB-. Now, depending upon the order among cys, trpA and trpB, the donor fragment will be cys+trpA-trpB+or cys+trpB+trpA-. Corresponding recipients will be cys-trpA+trpB- or cys-trpB- trpA+. In the first case where trpA is closer to cys, 4 crossover events will be required to get the prototroph, while 2 crossover events will be needed if trpB is close to cys. So, the sequence is cys-trpB-trpA.
1. In these studies always look for the supplement that is + for all the mutants. In this case G. So, G is the final metabolite. Then work backward, like supplement that gives ++ and so on.
Sequence is
5 4 2 1 3
E -------- A -------- C--------- B -------- D ---------- G
2. Here, either trpB or trp A is closer to cys
In the first case, recipient is cys+trpA-. So, it may be cys+ trpA- trpB+ or cys+trpB+ trpA- and incoming DNA fragment is cys- trpA+ trpB- or cys- trpB- trpA+. In either case a double crossover event will form prototroph (all +ve).
In the second case, recipient is cys-trpB-. Now, depending upon the order among cys, trpA and trpB, the donor fragment will be cys+trpA-trpB+or cys+trpB+trpA-. Corresponding recipients will be cys-trpA+trpB- or cys-trpB- trpA+. In the first case where trpA is closer to cys, 4 crossover events will be required to get the prototroph, while 2 crossover events will be needed if trpB is close to cys. So, the sequence is cys-trpB-trpA.
Friday, March 20, 2009
Practice problems.....Asignment 3
1. In a biosynthetic pathway, A to E are intermediate metabolites and final product is G. Several mutants are isolated that are defective in one or the other step in the pathway. The order in which these intermediates are formed is not known. Each intermediate is tested for its ability to support the growth to each mutant (1 to 5). In the following table, + indicates growth and – indicates no growth.

2. Two mutants at the tryptophan locus, trpA- and trpB-, are known to be close to a cysteine locus (cys). A bacterial strain of genotype cys+trpA-is conjugated to a donor strain that transfers cys-trpB-. The reciprocal cross in which donor transfers cys+trpA-to the strain that is cys-trpB-. In both the cases, the numbers of prototrophic recombinants are comparable. Determine the order of tryptophan loci relative to cysteine locus.

A)What is the order of A to E in the pathway?
B) At which point in the pathway is each mutant blocked?
[ hint: approach followed by Tatum and Beadle with Neurospora arg- auxotrophs]
B) At which point in the pathway is each mutant blocked?
[ hint: approach followed by Tatum and Beadle with Neurospora arg- auxotrophs]
2. Two mutants at the tryptophan locus, trpA- and trpB-, are known to be close to a cysteine locus (cys). A bacterial strain of genotype cys+trpA-is conjugated to a donor strain that transfers cys-trpB-. The reciprocal cross in which donor transfers cys+trpA-to the strain that is cys-trpB-. In both the cases, the numbers of prototrophic recombinants are comparable. Determine the order of tryptophan loci relative to cysteine locus.
{kind=link}
Thursday, March 5, 2009
Bacterial Conjugation
 Bacterial conjugation is the often regarded as the bacterial equivalent of sexual reproduction or mating; however it is not actually sexual as it does not involve the fusing of gametes and the creation of a zygote, it is merely the exchange of genetic information. In order to perform conjugation, one of the bacteria has to carry an F-plasmid, the other one must not.
Bacterial conjugation is the often regarded as the bacterial equivalent of sexual reproduction or mating; however it is not actually sexual as it does not involve the fusing of gametes and the creation of a zygote, it is merely the exchange of genetic information. In order to perform conjugation, one of the bacteria has to carry an F-plasmid, the other one must not.The F-plasmid (also called F-factor) is an episome (a plasmid that can integrate itself into the bacterial chromosome by genetic recombination) of about 100 kb (kilo base pairs) length. It carries its own origin of replication, called oriV. There can only be one copy of the F-plasmid in a bacterium (which is then called F-positive), either free or integrated.
Among other genetic information, the F-plasmid carries a tra and a trb locus, which together are about 33 kb long and consist of about 40 genes. The tra locus includes the pilin gene and controlling genes, which together form pili on the cell surface, polymeric proteins that can attach themselves to the surface of F-negative bacteria and initiate the mating. The pili themselves do not seem to be the structures through which the actual exchange of DNA takes place; rather, some proteins coded in the tra or trb loci seem to open a channel between the bacteria.
Wednesday, February 11, 2009
BEL204 assignment on Mendelian Genetics
1. Black wool sheep is due to a recessive allele b and white wool sheep due to dominant allele B. A white ram (male) is crossed to white ewe (female), both animals carrying the allele for black. They produce a white lamb that is then backcrossed to the female parent. What is the probability of the backcross offspring being black?
2. The shape of radishes may be long (SL SL), round (SLSR) or oval (SRSR). If long radishes are crossed to oval radishes and F1 then allowed to cross at random among themselves, what phenotypeic ratio is expected in the F2?
3. Plumage colour in mallard ducks is dependent upon a set of 3 alleles: MR for restricted mallard pattern, M for mallard and m for dusky mallard. The dominance hierarchy is MR>M>m. Determine the genotype and phenotype ratios expected in F1 from the following crosses: a) MRX MRX MR, b) MR MR X MR M, c) MR MX MR M, d) MRmXMm and e) MmXmm.
4. In Drosophila, ebony body colour is produced by a recessive gene a and wild type (gray) colour by its dominant allele a+. Vestigial wings by recessive gene vg and normal wing (wild type) by vg+. If wild type dihybrid flies are crossed and produce 272 progenies, how many of these are expected in each phenotype class?
Use comments to post your answers and do discussions on the assignment.
Tuesday, February 3, 2009
BEL102 Assignment
Tutorial problems (from Text 1, unless specified otherwise):
Problem set 1: 3.27, 3.32, 3.53, 4.23
Problem set 2: 4.13, 4.18, 4.30, 4.75
Problem set 3:
(a) In example 4.4-2, what is the degree of freedom for (i) overall process (ii) first extractor(iii) second extractor (iv) extract mixing point (v) distillation column?
(b) Learn how to calculate equilibrium composition from example 4.6-2. What is the limiting reactant and why?
Problem set 1: 3.27, 3.32, 3.53, 4.23
Problem set 2: 4.13, 4.18, 4.30, 4.75
Problem set 3:
(a) In example 4.4-2, what is the degree of freedom for (i) overall process (ii) first extractor(iii) second extractor (iv) extract mixing point (v) distillation column?
(b) Learn how to calculate equilibrium composition from example 4.6-2. What is the limiting reactant and why?
Monday, January 26, 2009
Mitosis
Mitosis is the process in which a eukaryotic cell separates the chromosomes in its cell nucleus, into two identical sets in two daughter  nuclei. It is generally followed immediately by cytokinesis, which divides the nuclei, cytoplasm, organelles and cell membrane into two daughter cells containing roughly equal shares of these cellular components. Mitosis and cytokinesis together define the mitotic (M) phase of the cell cycle - the division of the mother cell into two daughter cells, genetically identical to each other and to their parent cell.
nuclei. It is generally followed immediately by cytokinesis, which divides the nuclei, cytoplasm, organelles and cell membrane into two daughter cells containing roughly equal shares of these cellular components. Mitosis and cytokinesis together define the mitotic (M) phase of the cell cycle - the division of the mother cell into two daughter cells, genetically identical to each other and to their parent cell.
 Interphase
Interphase
The mitotic phase is a relatively short period of the cell cycle. It alternates with the much longer interphase, where the cell prepares itself for cell division. Interphase is therefore not part of mitosis. Interphase is divided into three phases, G1 (first gap), S (synthesis), and G2 (second gap). During all three phases, the cell grows by producing proteins and cytoplasmic organelles. However, chromosomes are replicated only during the S phase. Thus, a cell grows (G1), continues to grow as it duplicates its chromosomes (S), grows more and prepares for mitosis (G2), and divides (M).
Preprophase
In plant cells only, prophase is preceded by a pre-prophase stage. In highly vacuolated plant cells, the nucleus has to migrate into the center of the cell before mitosis can begin. This is achieved through the formation of a phragmosome, a transverse sheet of cytoplasm that bisects the cell along the future plane of cell division. In addition to phragmosome formation, preprophase is characterized by the formation of a ring of microtubules and actin filaments (called preprophase band) underneath the plasma membrane around the equatorial plane of the future mitotic spindle. This band marks the position where the cell will eventually divide. The cells of higher plants (such as the flowering plants) lack centrioles: with microtubules forming a spindle on the surface of the nucleus and then being organized into a spindle by the chromosomes themselves, after the nuclear membrane breaks down. The preprophase band disappears during nuclear envelope disassembly and spindle formation in prometaphase.
Prophase
{kind=link}
Prophase: The two round objects above the nucleus are the centrosomes. The chromatin has condensed.
Normally, the genetic material in the nucleus is in a loosely bundled coil called chromatin. At the onset of prophase, chromatin condenses together into a highly ordered structure called a chromosome. Since the genetic material has already been duplicated earlier in S phase, the replicated chromosomes have two sister chromatids, bound together at the centromere by the cohesion complex. Chromosomes are visible at high magnification through a light microscope.
Close to the nucleus are structures called centrosomes, which are made of a pair of centriole. The centrosome is the coordinating center for the cell's microtubules. A cell inherits a single centrosome at cell division, which replicates before a new mitosis begins, giving a pair of centrosomes. The two centrosomes nucleate microtubules (which may be thought of as cellular ropes or poles) to form the spindle by polymerizing soluble tubulin. Molecular motor proteins then push the centrosomes along these microtubules to opposite side of the cell. Although centrosomes help organize microtubule assembly, they are not essential for the formation of the spindle, since they are absent from plants, and centrosomes are not always used in meiosis.
Prometaphase
{kind=link}
Micrograph showing condensed chromosomes in blue and the mitotic spindle in green during prometaphase of mitosis
{kind=link}
Prometaphase: The nuclear membrane has degraded, and microtubules have invaded the nuclear space. These microtubules can attach to kinetochores or they can interact with opposing microtubules.
The nuclear envelope disassembles and microtubules invade the nuclear space. This is called open mitosis, and it occurs in most multicellular organisms. Fungi and some protists, such as algae or trichomonads, undergo a variation called closed mitosis where the spindle forms inside the nucleus or its microtubules are able to penetrate an intact nuclear envelope.
Each chromosome forms two kinetochores at the centromere, one attached at each chromatid. A kinetochore is a complex protein structure that is analogous to a ring for the microtubule hook; it is the point where microtubules attach themselves to the chromosome. Although the kinetochore structure and function are not fully understood, it is known that it contains some form of molecular motor. When a microtubule connects with the kinetochore, the motor activates, using energy from ATP to "crawl" up the tube toward the originating centrosome. This motor activity, coupled with polymerisation and depolymerisation of microtubules, provides the pulling force necessary to later separate the chromosome's two chromatids.
When the spindle grows to sufficient length, kinetochore microtubules begin searching for kinetochores to attach to. A number of nonkinetochore microtubules find and interact with corresponding nonkinetochore microtubules from the opposite centrosome to form the mitotic spindle. Prometaphase is sometimes considered part of prophase.
Metaphase
{kind=link}
A cell in late metaphase. All chromosomes (blue) but one have arrived at the metaphase plate.
{kind=link}
Metaphase: The chromosomes have aligned at the metaphase plate.
As microtubules find and attach to kinetochores in prometaphase, the centromeres of the chromosomes convene along the metaphase plate or equatorial plane, an imaginary line that is equidistant from the two centrosome poles. This even alignment is due to the counterbalance of the pulling powers generated by the opposing kinetochores, analogous to a tug-of-war between people of equal strength. In certain types of cells, chromosomes do not line up at the metaphase plate and instead move back and forth between the poles randomly, only roughly lining up along the midline. Metaphase comes from the Greek μετα meaning "after."
Because proper chromosome separation requires that every kinetochore be attached to a bundle of microtubules (spindle fibres), it is thought that unattached kinetochores generate a signal to prevent premature progression to anaphase without all chromosomes being aligned. The signal creates the mitotic spindle checkpoint.
Anaphase
{kind=link}
Early anaphase: Kinetochore microtubules shorten
When every kinetochore is attached to a cluster of microtubules and the chromosomes have lined up along the metaphase plate, the cell proceeds to anaphase (from the Greek ανα meaning “up,” “against,” “back,” or “re-”).
Two events then occur; First, the proteins that bind sister chromatids together are cleaved, allowing them to separate. These sister chromatids, which have now become distinct sister chromosomes, are pulled apart by shortening kinetochore microtubules and move toward the respective centrosomes to which they are attached. Next, the nonkinetochore microtubules elongate, pushing the centrosomes (and the set of chromosomes to which they are attached) apart to opposite ends of the cell. The force that causes the centrosomes to move towards the ends of the cell is still unknown, although there is a theory that suggests that the rapid assembly and breakdown of microtubules may cause this movement.
These two stages are sometimes called early and late anaphase. Early anaphase is usually defined as the separation of the sister chromatids, while late anaphase is the elongation of the microtubules and the microtubules being pulled farther apart. At the end of anaphase, the cell has succeeded in separating identical copies of the genetic material into two distinct populations.
Telophase
{kind=link}
Telophase: The decondensing chromosomes are surrounded by nuclear membranes. Note cytokinesis has already begun, the pinching is known as the cleavage furrow.
Telophase (from the Greek τελος meaning "end") is a reversal of prophase and prometaphase events. It "cleans up" the after effects of mitosis. At telophase, the nonkinetochore microtubules continue to lengthen, elongating the cell even more. Corresponding sister chromosomes attach at opposite ends of the cell. A new nuclear envelope, using fragments of the parent cell's nuclear membrane, forms around each set of separated sister chromosomes. Both sets of chromosomes, now surrounded by new nuclei, unfold back into chromatin. Mitosis is complete, but cell division is not yet complete.
Cytokinesis
Cytokinesis is often mistakenly thought to be the final part of telophase, however cytokinesis is a separate process that begins at the same time as telophase. Cytokinesis is technically not even a phase of mitosis, but rather a separate process, necessary for completing cell division. In animal cells, a cleavage furrow (pinch) containing a contractile ring develops where the metaphase plate used to be, pinching off the separated nuclei. In both animal and plant cells, cell division is also driven by vesicles derived from the Golgi apparatus, which move along microtubules to the middle of the cell. In plants this structure coalesces into a cell plate at the center of the phragmoplast and develops into a cell wall, separating the two nuclei. The phragmoplast is a microtubule structure typical for higher plants, whereas some green algae use a phycoplast microtubule array during cytokinesis. Each daughter cell has a complete copy of the genome of its parent cell. The end of cytokinesis marks the end of the M-phase.
The Transforming Principle

Griffith's experiment, conducted in 1928 by Frederick Griffith, was one of the first experiments suggesting that bacteria are capable of transferring genetic information through a process known as transformation.[1][2]
Griffith used two strains of Pneumococcus (which infects mice), a type III-S (smooth) and type II-R (rough) strain. The III-S strain covers itself with a polysaccharide capsule that protects it from the host's immune system, resulting in the death of the host, while the II-R strain doesn't have that protective capsule and is defeated by the host's immune system.
In this experiment, bacteria from the III-S strain were killed by heat, and their remains were added to II-R strain bacteria. While neither alone harmed the mice, the combination was able to kill its host. Griffith was also able to isolate both live II-R and live III-S strains of pneumococcus from the blood of these dead mice. Griffith concluded that the type II-R had been "transformed" into the lethal III-S strain by a "transforming principle" that was somehow part of the dead III-S strain bacteria.
Griffith used two strains of Pneumococcus (which infects mice), a type III-S (smooth) and type II-R (rough) strain. The III-S strain covers itself with a polysaccharide capsule that protects it from the host's immune system, resulting in the death of the host, while the II-R strain doesn't have that protective capsule and is defeated by the host's immune system.
In this experiment, bacteria from the III-S strain were killed by heat, and their remains were added to II-R strain bacteria. While neither alone harmed the mice, the combination was able to kill its host. Griffith was also able to isolate both live II-R and live III-S strains of pneumococcus from the blood of these dead mice. Griffith concluded that the type II-R had been "transformed" into the lethal III-S strain by a "transforming principle" that was somehow part of the dead III-S strain bacteria.
Experiment by Avery & MacLeod
 Today, we know that the "transforming principle" Griffith observed was the DNA of the III-S strain bacteria. While the bacteria had been killed, the DNA had survived the heating process and was taken up by the II-R strain bacteria. The III-S strain DNA contains the genes that form the protective polysaccharide capsule. Equipped with this gene, the former II-R strain bacteria were now protected from the host's immune system and could kill the host. The exact nature of the transforming principle (DNA) was verified in the experiments done by Avery, McLeod and McCarty and by Hershey and Chase.
Today, we know that the "transforming principle" Griffith observed was the DNA of the III-S strain bacteria. While the bacteria had been killed, the DNA had survived the heating process and was taken up by the II-R strain bacteria. The III-S strain DNA contains the genes that form the protective polysaccharide capsule. Equipped with this gene, the former II-R strain bacteria were now protected from the host's immune system and could kill the host. The exact nature of the transforming principle (DNA) was verified in the experiments done by Avery, McLeod and McCarty and by Hershey and Chase. Sunday, January 25, 2009
Use it!!!!!!!!!!!!
hey guys nd gals...........use this place to post your doubts and any of the information u like to share...........for the doubts (academic or non-academic) other users shall answer dem to your benefits.............
Thursday, January 15, 2009
BEL102 Assignment
Assignments (from Second Edition of Text 1, unless specified otherwise):
- Chapter 2, problems 7, 8, 9, 10, 11, 14, 18, 27, 31, 40.
Subscribe to:
Comments (Atom)